









Decoding the song of the pied butcherbird: an initial survey
“Of course viewing culture as something which originates in a natural function, and imagining that it turned out to bring a new end beyond pure survival, may look heretical both to a large majority of biologists and to many musicians as well… I can only say, as a composer, that Cracticus nigrogularis, the pied butcherbird, is a kind of colleague.”
François-Bernard Mâche (2000: 479)
Abstract
Definitions of music invoking the human cutoff point are reviewed. The pied butcherbird Cracticus nigrogularis is suggested for a zoömusicological case study on how birdsong might be like the human animal’s music (whether homologous or analogous). Portamento as an impediment to “off-the-shelf” musicology in the case of birdsong analysis is discussed. It is proposed that the employment of different types and levels of description could facilitate the most fecund survey and analysis. An initial survey on how pied butcherbirds use notes, calls, and song is presented, including sonograms and standard music notation, followed by an elucidation of a repertoire of procedures found in both “music” and pied butcherbird song. It is concluded that their elaborate song culture seems to overreach biological necessity, indicating an aesthetic appreciation of sound is present in the pied butcherbird.
Key words: zoömusicology, birdsong, pied butcherbird, portamento, sonogram, notation.
The search for a simple declarative sentence to pin down the moving target of music in the twentieth century and beyond, along with the quest for a succinct list of universals in music, grants particular appeal to the human cutoff point in a definition of music. Music as a uniquely human activity is a recurring theme in the literature—witness Merriam’s “Music sound cannot be produced except by people for other people” (1964: 6); Sessions’ “music is created by human beings” (1950/1974: 11); Kolinski’s “Music has been created by man” (1967: 1); Harrison’s “non-linguistic sound, when used (with some degree of intention) by human beings” (1977: 30); Blacking’s “humanly organized sound” (1995: 10); Bowman’s “a product of human minds” (1998: 69); Cook’s “humanly generated sounds” (1998: 4); and Cross’ “a peculiarly human phenomenon… quite outside the repertoire of behaviors of other species” (2003: 109-110). List frames ethnomusicology as “the study of humanly produced patterns of sound,” and thus “bird song lies without the province…” (1979: 1). Kivy attacks the issue obliquely, invoking the pseudo-linguistic model:
For to say we hear bird songs as if they had syntactical properties is not to ascribe syntactical properties to them, any more than we are describing a monster when we say of someone that it is as if he had eyes in the back of his head. However, as soon as we take being able to hear bird noises as music to imply that therefore they are music, we are saying that they literally have syntactic properties; and that is a conceptual impossibility. A natural object cannot, as a matter of logic, have syntactic properties, whether it is a bird’s “song” or anything else… (Kivy 1990: 24-25).
Equating object with an animal is problematical. Are songbirds mere curiosities within the landscape, interchangeable with a children’s bird whistle or a music box? More to the point, biologists regularly remark on the coding rules of syntax in animal vocalisations (Richman 1987: 201; Bradbury and Vehrencamp 1998: 461, 494; Doupe and Kuhl 1999: 571: Rogers and Kaplan 2000: 86). Birdsong is not haphazard, “random handfuls of notes” (Hartshorne 1958: 422)—but neither is it a language. In fact, music is not a language. It has “no evident, immediate fixed consensual reference” (Stevens 2004: 25). Mâche, who coined the word zoömusicology in 1983 (1983/1997: 95-160), warns of the classic syllogism: “Language distinguishes man, and music is a kind of language, therefore music is a purely human cultural fact” (ibid.: 73).
Martinelli imagines zoömusicology as the study of the “aesthetic use of sounds among animals” (2001: 3); such is the scope of the birdsong study herein. For the purpose of my investigation, it is not necessary that birdsong cross that final hurdle and be classified “music.” (Having avoided the yoke of Merriam’s “White Knight Concept,” I do nonetheless admit to his “Duty of Preservation Concept” (1963: 207), “music” or not.) For those inclined towards an aesthetic appreciation of birdsong as music, including perhaps the birds themselves, the connection seems obvious. For others, it is a line that cannot be crossed (until, perhaps, inter alia we have a theory of mind for animals[i]).
A bird’s song can function as legal deed to his territory (a national anthem[ii])—an auditory “keep out” sign—or as a “come hither” (a serenade) to his female counterpart. It can serve as a group password (Fitch 2006: 186). In fact, “Why do birds sing?” involves a complex web of many correct answers (ibid.: 174) and as many unanswered questions (Rothenberg 2005).[iii] Although not intended for incidental human eavesdroppers, through the centuries birdsong has inspired artists of all stripes, as well as philosophers and just plain folk; occasionally we even propose our own imagined translation of their song or “mood.” The melodic inventions of birds have been appropriated by composers, but by-and-large musicians have failed to pursue serious interrogations into the musical life of birds. At the point when technology arrived that would assist us to notate the notoriously difficult songs of birds, we musicians defaulted. The advent of the tape recorder, followed by further technological advances such as digital recording and computer analysis programs, allowed biologists to leapfrog over musicians and assume the mantle in the study of avian acoustics and its concomitant behaviour. A groundswell of activity followed.
Vocal learning is rare. Aside from the human animal, evidence for it is confirmed only in oscine songbirds, parrots, and hummingbirds, while circumstantial evidence for it exists among some marine mammals and bats (Wilbrecht and Nottebohm 2003: 135). The predisposition for perception and learning evolved separately three times in birds (Doupe and Kuhl 1999: 573).
It was clear to Darwin, and has remained unargued ever since, that bird song is analogous, not homologous, to human song (our common ancestor, a Paleozoic reptile, did not sing), and the same can be said for whale and seal song (Fitch 2006: 183).
However, Martinelli claims that a concept for music exists in non-human animals and thus the “sound manifestations in non-human animals are homologous to musical manifestations in humans. They are not simply analogous” (2002: 106-107). Whatever the vote on “same” or “different” when applied to origin and function, clearly all agree that birdsong is “like” the human animal’s music in that it involves learning.
The process of song acquisition for songbirds follows a trajectory from the first tentative and poorly structured notes (subsong), to the intermediate stages (called “plastic song” by biologists because although the notes are more structured, they are still unstable and highly variable), to the mature adult stage when stereotypy is achieved (“crystallized song”) (Thorpe 1958; Thorpe and Pilcher 1958; Marler and Peters 1982; Hultsch and Todt 2004: 80-107). Learning is accomplished via cultural transmission and can be vertical (learning from parents), horizontal (learning from members of the same generation), and oblique (learning from unrelated birds of different generations) (Lynch et al 1989: 634; Baptista and Gaunt 1997: 24-25).
Compared to songs, birdcalls tend to be shorter, simpler, and innate. The word “innate” recalls the dichotomous thinking of the nature/nurture debate, over which much ink has been spilled. Ornithologists now rely less on the words “learned” and “innate,” which imply strict boundaries, and have gradually replaced them with concepts such as an “inherited tendency” (Thorpe 1958: 557), “instincts for inventiveness” (Marler 1994: 63), “learning preferences” (Marler 1997: 503), “song templates” (ibid.: 503), and “auditory templates” (Adret 2004: 306). Nature or nurture has evolved into nature and nurture, complementary activities often now viewed as a continuum from biology to culture.
Once establishing the basic song learning cycle, biologists went on to refine their knowledge. Space does not permit a further examination of this subject, nor does this writer intend to cite studies that depended on experiments such as the deafening of hatchlings, the separation of mated pairs, the playback of taped conspecific song, and the “harvesting” of bird brains for neurological research, where songbirds serve as stand-in humans for researchers. Suffice it to say that much remains to be discovered and very few species have been studied in depth, so that much of what we think we do know is based on several “white rats” of the bird world (Baptista 1975: 1). With a few notable exceptions (Craig 1943; Sotavalta 1956; Armstrong 1973; Baptista and Keister 2005), the preoccupations of most ornithologists concern biological and evolutionary questions (the ontogeny and function of song, for example, as one would expect), rather than musical ones. However, one does find numerous sidelong glances towards the music camp—the comment that research into song complexity “leaves us to puzzle over the resulting richness and variety…” (Catchpole and Slater 1995: 191) and the description of birdsong improvisation as “assuaging an appetite for novelty” (Marler 1981: 92) are cases in point. The scientific attraction of birds is that they are easier to study than a number of other species. However, my interest is not in setting up experiments but in observing what birds spontaneously produce with a minimum of intrusion.
The pied butcherbird Cracticus nigrogularis is a medium-sized songbird with sharply contrasted black and white plumage and a black hood and bib. The species is non-migratory, and its territory includes much of mainland Australia, with a notable absence from the southern coast and desert areas (Higgins, Peter, and Cowling 2006: 516). Males and females are generally indistinguishable in appearance. The (assumed) male sings spring song in the pre-dawn hours and during moonlit nights. Both sexes participate in the dawn chorus, and their daytime vocalisations, often antiphonal duos or trios, can be heard off and on throughout the day. Their rich, diversified voice produces mellow sounds; the metaphor of a piping flute, a cornet, or an organ is often noted, as indicated by one of the bird’s alternative names, “organ-bird.” Magnificent, wonderful, glorious, superb, delightful, beautiful, melodious, bubbling, best, finest, king of songsters—the list of superlatives describing the voice is considerable. Hartshorne described it as “the true ‘magic flute’, the perfection of musical tonality coming from a bird” (1953: 118). They are accomplished mimics. Their song impressed Olivier Messiaen, who incorporated pied butcherbird song into his last work, Éclairs sur l’au-delà:
J’ai écouté très longuement ces chant d’oiseaux Australiens, et j’ai même essayé de les noter. Ils sont tous d’un grand intérêt, specialement Pied Butcherbird et Grey Butcherbird, qui sont marveilleux.[iv]
Surprisingly, despite their wide distribution and noted singing ability, no detailed studies have been undertaken of their song or their behavior, save one thesis on breeding.

Fig. 1. A pied butcherbird featured in a warning poster used by the New South Wales National Parks and Wildlife Service during spring nesting season when birds can become aggressive. (Used with permission from DECC (Department of Environment and Climate Change), undated. PO Box A290 Sydney South NSW 1232.)
I first encountered the pied butcherbird five years ago in the outback of Western Australia. As a violinist/composer trained in the Western classical music tradition, I have spent much of the last 35 years transcribing music of the American vernacular, from fiddle tunes to jazz, from country to Cajun. Residencies in the Dominican Republic and Budapest saw me extend my research to music of the Caribbean and eastern European folk music. While some of these styles I actively performed in, more often the purpose of my research was to expand my “toolkit” as a composer and improviser. My expertise lay in transcription.
I determined to compile the song phrases of the pied butcherbird, a sort of composer’s compendium of natural forms for my use and others. I wanted to examine the musicality of their songs and in the process, if not take the field of birdsong study back for musicians, at least to participate in the discussion.[v] Of course, the avian voice is more than a commodity awaiting full realisation by the composer. My instincts suggested I would develop this composers’ catalogue not to improve on pied butcherbird song, nor to mimic it, but to immerse myself in their musical landscape and emerge with a substantive knowledge of the acoustical constructs of another species with whom we share this planet.
The Australian composer and music critic Henry Tate had prefigured this birdsong notebook.
The use of bird calls as the basis for evolving new musical scales was a theme to which [Henry Tate] returned again and again. One of his most specific proposals was for the cultivation of a scale rationalized from the calls of the butcher bird, a bird which, despite a perhaps unprepossessing name, is certainly the equal in musical eloquence and beauty of such celebrated singing birds as lark and nightingale. Tate felt that a scale abstracted from the musical language of the butcher bird could have a flattened second, a major third and a minor sixth. He perceived not only the modal affinities of this scale but also its relation to the contemporary practice of Bartók—an act of perception which, in 1922 (the date of the essay in which the reference occurs), put him approximately thirty years ahead of the majority of his fellow musicians. Whether or not Tate reproduced the calls of the butcher bird accurately is open to doubt: he admitted that the flattened sixth he proposed for this scale was his own notion of what would be in sympathy with the butcher bird’s flattened second. But details of this kind are less important than the practice that might grow from such an observation (Covell 1967: 104-105).
This suggested the task at hand: to notate their song phrases and then proceed to decode their underlying principles of organization, confirming or correcting Tate’s deflected scale.
The danger in any convention rests in its ability not to seem like one. Pitch (and its concomitant feature, frequency) was by default the most significant quantifiable characteristic as well as the point for insight into musical structure. (Ethnomusicology and musicology are filled with studies that would collapse without a pitch reference.) Pied butcherbird song is largely melodic in nature and often delivered in a slow and regular way, suggesting a straightforward path. Microtones outside the equal-tempered scale were expected, but the intention was to limit notation to the pitches available on the staff, supplemented by arrows placed atop noteheads pointing up or down to indicate those occasional microtonal pitches that the ear, which is generous in matters of pitch discrimination (Seashore 1938/1967), would not quantize into 12tET (twelve-tone equal temperament). The goal was to produce readable, non-specialised notation for my monograph: Notes from the Songbook of the Pied Butcherbird.
Ornithology is perhaps the only science where amateurs contribute serious and respected work; wildlife recordists are a case in point. Many of them shared their pied butcherbird field recordings, paired with notes containing the species name, date, time, location, habitat description, temperature, behavioral context, number of individuals, distance to the bird, and type of recording equipment. For the fieldwork, a Sennheiser ME67 shotgun microphone, a Sony portable high definition minidisc recorder, and a GPS indicator to note exact orientation were employed.
Although this writer possesses absolute pitch, the advice was to double-check all transcriptions with software, so the results would be “scientifically” reliable. Sonagraphic analysis was carried out on a personal computer using the software Amadeus II, version 3.8.7[vi]. Sometimes called a sound spectrogram, this graphic representation of sound consists of an x-axis depicting time and a y-axis for frequency, while a grey-scale denotes relative amplitude. Amadeus’s vertical frequency axis displays no numbers; however, by clicking anywhere in the sonogram, a pop-up window appears, showing the frequency corresponding to the location of the mouse, as well as the note that is closest to that frequency and the standard reading for that note: e.g. 1313.53 Hz, E6 (1318.51 Hz). Sonograms were printed using the software Raven 1.2[vii].
As an adjunct to the project, one scientist suggested the taking of frequency measurements to examine if the birds sing in simple integer ratios, something approaching just intonation. It was assumed that for the study to be taken seriously by scientists, quantification was a key strategy. Considerable time was spent trying to sort out how to best detect “exact” frequency (Taylor 2006). Obviously, a human element is involved in clicking in a sonogram window or otherwise choosing the area to be measured; the results were not entirely consistent. Other software, whether by indicating the area of the most intense signal or requiring one to draw a square around the note in question, also produced inconsistent numbers on multiple measurements of the same material.
In the process of surveying the software best capable of consistently measuring frequency, another obstacle surfaced. Although pied butcherbird song seems to consist largely of discrete pitches, closer examination revealed that that is rarely the case. Portamento is omnipresent. Sometimes termed a “frequency-modulated tone” or “slur” by biologists, portamento is also often mistakenly called “glissando” (which implies production on an instrument with fixed semitones, such as the piano or harp, while “portamento” does not distinguish the intervening notes in its glide (Boyden and Stowell 2007).
Those cultures whose music relies upon pitch bends, portamenti, microtones, blue notes, and arabesques of ornament and decoration may find the fixed pitches of Western art music’s equal-tempered system limiting. While pitch glides and microtonal strategies could “work” for an audience member from any culture, this musicologist hoped to come across a majority of discrete pitches for more reasons than ease of melodic notation.[viii] The relational aspects of pitch that ensue after measuring individual notes, from the simplest inter-tone relationships to the hierarchical relationship among the tones such as intervals, scales, melodic contour[ix], tonality, structure—these key components of the typological research I had imagined appeared to be slipping away. I wrote a somewhat forlorn letter to François-Bernard Mâche, who responded:
You have reason to first trust your ear, then measurements. For complex sounds, like most of those of birds, our ear generally defines a subjective pitch that sonograms do not reveal, particularly when there are portamenti. The exact transcription is unusable for a musician, and the subjective transcription is unusable for a biologist. This is one of the ambiguities of our work. I believe that it is best to first define the goal of the analysis, which will then determine the type and degree of simplification of the acoustic terrain. What is pertinent for the musician will not always be so for the acoustician, and what the ear picks up on is not always present in a sonogram. I used to make my transcriptions too precise, rendering them nearly illegible to others. These days, I simplify.[x] [HT trans.]
Fearful of breaching the canon of scientific objectivity, my earnestness had allowed me to slip into the netherworld of an “unmarked category.” I needed to come out and be a musician, acknowledge my trained ear, and turn it towards pied butcherbird song, and “I had to learn to resist the easy solutions my tonal theory training had given me” (McClary 2000: x).
Science excels in the telling; music suggests.
Nor must we lightly accept the damaging distinction between pure and applied science, between use and abuse of science, and even between nature and culture. All are versions of the philosophy of science that exploits the rupture between subject and object to justify the double ideology of firm scientific objectivity and mere personal subjectivity (Haraway 1991: 2).
“Facts” can be deficient. Science’s instruments, software, and experiments enumerate and explicate, but we know from physics that many aspects can neither be definitively tested nor accounted for; that does not mean they do not exist. I became determined to confront musical issues and not technological ones. I had survived the “lure of numeracy” (Gourlay 1978: 26). I felt sure that a musician, one well-trained and with a broad experience of music, has essential insights to offer biologists.
Game theory tells us that the best approach is to have a mixture or range of strategies from which to choose, some perhaps more often than others, rather than a single strategy (Siegfried 2006). After the first blind alley, seeking out several concurrent paths of description seemed sensible, with the concern that pursuing only one way of knowing could not only result in another blind alley but could also create an imbalance. Multiple ways of knowing were required for the fullest picture possible. I was now nudging things along, operating on guesswork—hopefully, the inspired guess. At least one scientist also embraced this modus operandi.
The English, who have developed their government in this direction, call it “muddling through,” and although a rather silly, stupid sounding thing, it is the most scientific way of progressing. To decide upon the answer is not scientific. In order to make progress, one must leave the door to the unknown ajar… (Feynmann 1999: 115).
In considering how to extend the graphic potential of standard music notation without making it overly complicated, and understanding that all descriptions are partial, hybrid notations were avoided. The music staff was paired with a sonogram. “The only really true notations are the sound-tracks on the record itself,” Bartók suggested (Bartók and Lord 1951: 3). However, a recording is not a tangible fact, nor is a sonogram. Technology can help us with things that we grasp intuitively, and the trained ear is a powerful partner (List 1974: 375). Elsewhere, Bartók observed that “… having the best material equipment is not enough: the equivalent intellectual equipment is just as important” (Suchoff 1976: 10).
In matters of uncertainty, whether pitch or rhythm, the deference was to the human ear for the final decision. Amadeus allows for playback at varying speeds while maintaining the original pitch. Half-speed was occasionally employed, but rarely was anything of merit not already observed in the sonogram and heard at original speed discovered. Notation was accomplished with the music notation software Finale 2007[xi]; its playback feature was often drawn on to confirm the notation. At times the same phrase was notated twice, one more “bare bones” than the other. The most challenging notational aspect was portamento. Normally, either the beginning or end of the glide was heard as the resultant pitch; a normal notehead indicates this, while a grace note serves for the other end of the glide. No attempt was made to measure the duration of the glide, as the speed is usually continuous. Each phrase was set into one bar; thin double bar lines marked out phrases and indicated a momentary stop in singing.
Every effort was made to employ basic terms and simple definitions to facilitate communication among readers with diverse backgrounds. There has been a notable lack of consensus among biologists. Shiovitz (1975: 133) counted five terms for “phrase,” six for “note,” and in total twenty different terms employed to describe eight “song” units, while suggesting standard terms. Nearly two decades later one study reviewed over 80 definitions of “birdsong”, finding little agreement in its definition or how to differentiate songs from calls (Spector 1994), while another called for unanimity of method as it tallied up 28 song unit identifications (Thompson, LeDoux, and Moody 1994). One looks back to the sage advice of Craig decades prior, that while no ornithologist had framed a satisfactory definition of birdsong, we all know a song when we hear one (1943: 169). A biologist himself, he went on to proclaim:
One reason why biologists have never successfully defined bird song is that it is not a purely biological concept. In order to develop an adequate concept of bird song, we must make progress not only in ornithology but also in musical esthetics (ibid.: 172).
For our purposes, pitch is employed to describe what is heard; frequency expresses what is measured. A note is defined as a discrete sound unit, whether modulated or not, and is represented by a continuous trace on the sonogram. (Harmonic overtones appearing as stripes above the fundamental are considered as part of the fundamental.) A motif is a coherent subsection of a phrase. Phrases are a recognizable and orderly group of notes and are separated by pauses, which are generally of the order of several seconds. A song is a sustained singing performance.
A bird’s production mechanics differ significantly from that of human animals. Although birds possess a larynx at the top of their trachea, their vocal mechanism is the syrinx (Fletcher 1992: 317), consisting of a valve in each bronchi just below the junction with the trachea. The singing of two unrelated notes is possible (Greenewalt 1968: 179; Nottebohm 1971: 229), although most birds either double a note with each valve, use one valve for high notes and the other for low notes, or fail to use one of the valves (Fletcher 2006: 35). How the syrinx functions is a complex matter under continuous review; many issues remain unresolved (Gaunt 1983; Nowicki and Marler, 1988; Catchpole and Slater 1995: 22; Goller and Larsen 1997, 2002). Here, then, are some of the sounds that pied butcherbirds can produce and that interest them and me.
1. NOTE STRUCTURE
Fig. 2 illustrates their basic note types (after Thorpe and Lade 1961: 236-244). Due to the variability of field recordings (dating back as far as forty years) from multiple recordists and a range of equipment, amplitude (volume) could not be reliably assessed. Usually it was possible, nonetheless, to determine whether the sound signal was strong, moderate, or weak. These were noted in the transcriptions, along with accent signs if a note stood out for its intensity and attack.
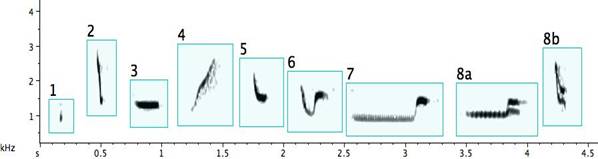

Fig. 2. Exemplars of basic note types of the pied butcherbird: 1. a very short note (less than .1sec) within a narrow frequency span (Extremely short notes sound click-like, whatever their frequency.); 2. a very short note covering a wide, but not simultaneous, frequency span; 3. a note with an almost constant frequency; 4. a note with an upward inflection; 5. a note with a downward inflection; 6. a warbling note; 7. two or more notes joined together by a tail; 8a/8b. simultaneously produced notes known to emit from one bird; 9. complex, “buzzy,” or “noisy” notes (where the energy is distributed simultaneously at multiple frequencies).
The pied butcherbird builds signal diversity with short repeated notes of various types, as in Fig. 3. Several deserve detailed description: Example 1: A trill (an alternation of two different, though near, pitches) (Broughton 1963: 12). A trill is rare in pied butcherbird song; this is the sole example I encountered. The bird was singing at the same time as—and one is tempted to say “along with”—a flute concerto played regularly on the stereo at the recordist’s home and clearly audible outdoors. Example 2: A rattle (a repetitive sequence of short pulses on the same or similar pitch) and marked on my music notation with “R” above the note. (When it does not deviate in pitch, it is equivalent to a tremolo.) Example 3: A quasi-rattle (the pulses fail to completely separate to the ear and on the sonogram) and marked on my music notation with “QR” above the note. Examples 12 and 13: “Noisy” rattles with a wider than normal frequency range. Example 14: A long rattle, in this case at a rate of 25 pulses spanning 1.5sec.
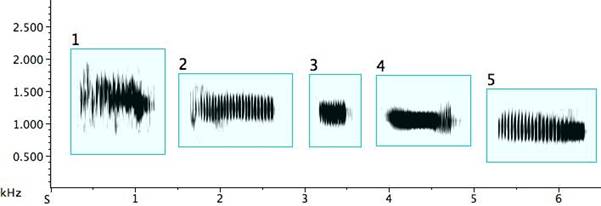
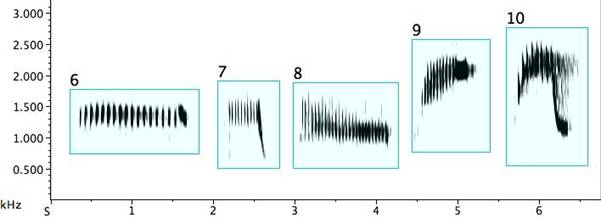
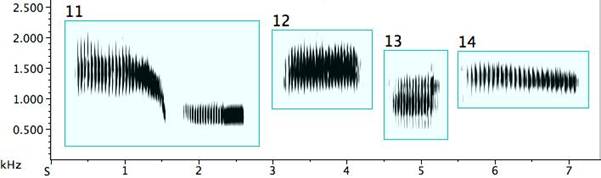
Fig. 3. Exemplars of short repeated notes: 1. a trill; 2. a rattle; 3. a quasi-rattle; 4. a note becoming a rattle; 5. a rattle becoming a note; 6/7. rattles ending with a flourish; 8. a descending rattle; 9. an ascending rattle; 10. an ascending and descending rattle; 11. two rattle types from one bird; 12/13. “noisy” rattles; 14. a long rattle.
Some notes call to mind mnemonic catchwords, such as the listing below of “blip”, “blop”, and the like. The times for the examples detailed in Table 1 and drawn in Fig. 4 could vary while the mnemonic device might still apply. The results are highly subjective, part acoustics and part psychoacoustics, and perhaps their key value is to indicate a change in timbre, even if they fail to adequately convey the sound quality.
|
Table 1: Mnemonic catchwords |
|
1.“Blip”: a relatively short (.1sec), descending (1141 Hz ¯ 1012 Hz) note. |
|
2.“Blop”: a relatively short (.07sec), descending (1249 Hz ¯ 732 Hz) note (“Blop” sounds lower than “Blip.”). |
|
3.“Tok”: a short (.035sec), hollow sounding note (970 Hz ¯ 710 Hz) (“Tok” sounds are either relatively stable in pitch or descend slightly.). |
|
4.Several reiterations of “Tok” (.065sec, 861 Hz) followed by a group of hollow sounding but higher “Chook” (Australian onomatopoetic vernacular for “chicken”) (.05sec, 1206 Hz). |
|
5.“Chook” (.04sec, 1184 Hz). |
|
6.A note of almost constant pitch followed by “Tok” (.03sec, 1249 Hz). |
|
7.Several reiterations of “Tok” (.02 ¯ 04sec, 1141 Hz) resolving to a note of almost constant pitch. |
|
8.Several reiterations of “Chook” (.04sec, 1120 Hz) followed by a single, higher, descending “Chip” (.04sec, 1637 ¯ 1314 Hz). |
|
9.The first of four “Chip” sounds (.03sec, 2110 ¯ 1615 Hz). These descending notes tend to be shorter and higher than the “Blip-Blop” family; they are not hollow-sounding like “Tok” nor are they of almost constant pitch like “Chook.”). |
|
10.“Chip” (.03sec, 2239 ¯ 1572 Hz). |
|
11.“Chip” (.03sec, 2347 ¯ 1960 Hz). |
|
12.“Chip” (.03sec, 2218 ¯ 1572 Hz). |
|
13.A double “Chip” (.06sec, 2638 ¯ 1421 Hz). Although slower than the other “Chip” sounds, perhaps because the frequency change is steeper, this pair maintains a “Chip” sound rather than the slower “Blip-Blop” family. |
|
14.The first of three “Wow” sounds (.15sec, 1098 ¯ 1421 ¯ 969 Hz). These short notes rise and then fall to a lower point than the initial pitch. |
|
15.“Wow” (.13sec, 1572 ¯ 1820 ¯ 1087 Hz). |
|
16.“Wow” (.24sec, 1044 ¯ 1151 ¯ 743 Hz). |
|
17.“Woop”: a relatively short (.05 ¯ .07sec), ascending note (689 ¯ 1034 Hz) usually repeated and here delivered at eight per second. |
|
18.By the time “Woop” is delivered at 13 per second, it begins to take on the timbre of an electronic, rather than an acoustic, signal (.03sec, 775 ¯ 980 Hz). |
|
19.An abrupt, downward, linear frequency sweep (marked “ES” in my notation for the electronic signal it suggests, and known in computer jargon as “pitch bend”). The faster the delivery or the steeper the frequency decline, the more heightened the effect. This example is delivered at twelve per second on the interval of a minor sixth (.03sec, 1324 ¯ 807 Hz). |
|
20.This example is delivered at a faster rate but covers a narrower frequency range, a perfect fourth (.04sec, 1604 ¯ 1141 Hz on average). |
|
21.This example travels two octaves (.075sec, 2563 ¯ 657 Hz). |
|
22.This example begins with a note of almost constant frequency that is interrupted by three descending notes (all from one bird). The second of the three travels the interval of a major ninth (.06sec, 1905 ¯ 851 Hz). |

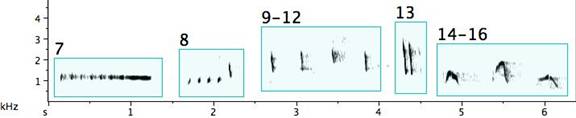
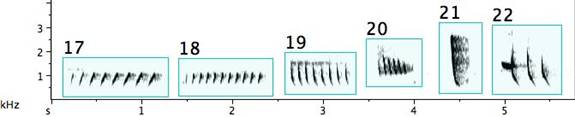
Fig. 4. Exemplars of notes suggesting mnemonic catchwords or electronic-sounding signals: 1. “Blip”; 2. “Blop”; 3. “Tok”; 4. several reiterations of “Tok” followed by “Chook”; 5. “Chook”; 6. a note of almost constant pitch followed by a “Tok”; 7. several reiterations of “Tok” resolving to a note of almost constant pitch; 8. several reiterations of “Chook” followed by a “Chip”; 9 ¯ 12. four different “Chips”; 13. a double “Chip”; 14 ¯ 16. “Wow”; 17/18. “Woop”; 19 ¯ 21. abrupt, downward, linear frequency sweeps (or portamenti); 22. a note of almost constant frequency interrupted by three extreme portamentos.
One questions why the pied butcherbird devotes effort in crafting notes that are so acoustically complex. What advantage might there be to the bird? What might the relationship be, if any, between complexity and reward? What might be the nature of the reward?
2. CALLS
Call notes are associated with the general (non-sexual and non-territorial) maintenance activities of feeding, flocking, migration, and response to predators (Thorpe 1964: 740). Fig. 5 presents the everyday calls of the pied butcherbird, as detailed in Table 2.
|
Table 2: Calls |
|
1.A food begging call from a nestling. |
|
2.A food begging call from an adult.[xii] |
|
3.A strident “reek” alarm call. Note the stripes of harmonics. Pied butcherbirds emit broad frequency alarm calls that are termed “nasal” (harmonic) rather than “harsh” (noisy) (Jurisevic and Sanderson 1994: 71). |
|
4.A scolding call given to a cat. This would fall under the rubric of agonistic signals (Bradbury and Vehrencamp 1998: 359). |
|
5.Sometimes birds deliver modulated, non-vocal mechanical sounds (Prum 1998: 977). These two beak claps delivered in quick succession, given as a sign of aggression to an Australian magpie, are not technically a call. The literature does not recount, nor do present recordings indicate, that this species makes use of other mechanical sounds, such as wing feather specialisations or drumming with beaks or tools. |
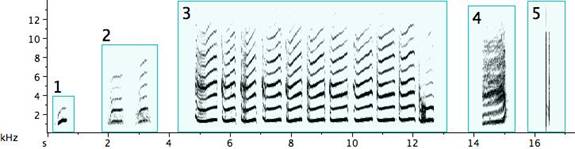
Fig. 5. Exemplars of call notes: 1. a food begging call from a nestling; 2. a food begging call from an adult; 3. a strident “reek” alarm call; 4. a scolding call; 5. two beak claps.
A number of other contexts exist where the pied butcherbird could call. In most cases, the call of choice is presented in Fig. 6. Various informants identified it as a flight call, a contact call, a separation call, an advertising call, a dominance call, a mutual recognition call, a mobbing call, and an “I am a pied butcherbird”[xiii] call. It is diagnostic for the species, meaning that one can identify the bird by this, whether or not it has been sighted. Because there is no common motivational basis for this call, but instead it accompanies various behaviours, the term “species” call was coined (Taylor 2005). Like the other calls, this one is generally stereotyped across the continent. Depending on how many notes it is delivered in, it has been referred to with the mnemonic “8-2-2” (three notes) or “eight twenty-two” and “por-rk-it-tee” (four notes). Occasionally it is extended to five notes. The notes often dip and rise and are usually delivered within the F7 (2794 Hz) ¯ G#7 (3322 Hz) range[xiv], an octave above the standard singing register.
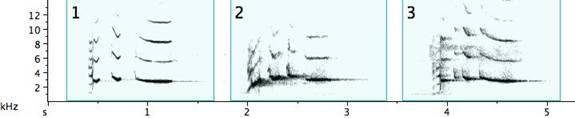
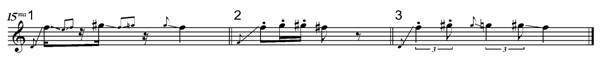
Fig. 6. Exemplars of species calls from three different birds (top) paired with music notation (bottom).
3. CALLS IN SONG
The differences between calls and songs are usually portrayed as clear-cut (Marler 2004: 32; Hall-Craggs 1961: 367). However, some find the distinctions not so easily demarcated (Johnson 2003: i; Smith 1991: 250); my research would support this latter view. Pied butcherbirds lack a species-typical song. The musical coordinates of their song are rather like Australian Aboriginal songlines, phrases overlapping and gradually changing as they spread out across the continent. In the 175 hours of recordings examined from 260 locations, no song phrases are repeated in more than the immediate and nearby territories they were found in, excepting two. Both of these appear to be re-workings of the species call.
When two or more birds are present and singing, the species call might be used by one rather like the interjection “Amen” in an American Southern Baptist church meeting. Delivered in apparent random fashion, it hovers in a border territory between call and song. Other times, the species “call” is delivered in antiphonal song where it seems a coherent part of the musical statement, a call-become-song (Fig. 7: S1). The “call” is also frequently found in solo song (Fig. 7: S2). The birds possess a repertoire of techniques for varying the motif. Unlike the pitch stability found in the call proper, when delivered as a motif in either antiphonal or solo song, the frequency of the “call” is no longer stereotyped. Nevertheless, the frequency does shift upwards significantly for a falsetto effect.
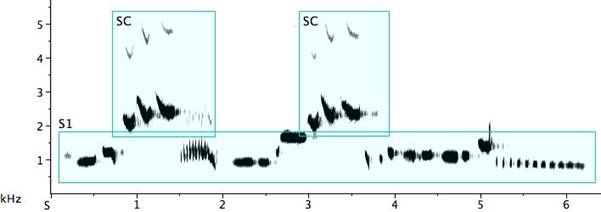

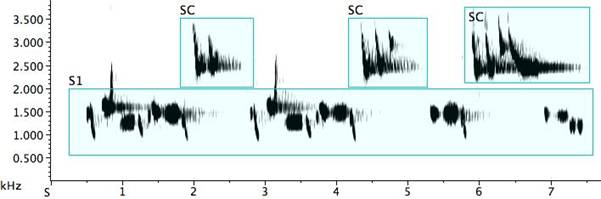
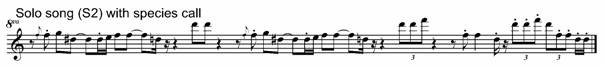
Fig. 7. Exemplars of species call notes incorporated in song: 1. antiphonal song #1 (S1) (R=rattle) with species call (SC) (the highest lines on the sonogram are harmonic overtones rather than whistled octaves); 2. solo song (S2) with species call (SC).
The abundant portamenti in the solo song above are not notated in the interest of simplicity. Note the octave jump from D6 to D7, the additive treatment of the species call motif, and D-D-F inversion to F-F-D (also an octave in distance). While the species call motif is altered on each of three deliveries, the other song phrase with which it is paired is repeated exactly the second time, delivered in truncated form the third time, and after the final species call motif, we have an end flourish of this same truncated motif with the final note doubled.
This call is commonly subjected to another treatment to build singing performances. Here the innate elements of the call are revised down an octave and the tempo greatly reduced. The motif is articulated short-long or short-short-long. As with the call, this “short-long” motif is normally delivered on stereotypical frequencies. Fig. 8, as explicated in Table 3, explores variations on a theme.
|
Table 3: Variations on a the short-long motif |
|
1.A descending half step. |
|
2.An ascending half step. |
|
3.A rhythmical variation on measure one. |
|
4.A descending whole step, followed by a motif reminiscent of the species call in that it jumps a large interval and stays in that register (rather than utilising a gap-fill strategy) with a triplet composed of semitones. |
|
5.Descending by a whole step, then a half step. |
|
6.A variation on measure five involving deflection. |
|
7.Similar to measure six, with the entry point a major second lower. |
|
8.A fully developed phrase built upon the motif (G-F on beats two and three), outlining a G dominant seven chord. |

Fig. 8. Exemplars of the “short-long” motif. (QR=quasi-rattle.)
One bird presents the phrase with an ultra-noisy rattle at the end—an aggressive “prew” sound finds the pied butcherbird acting momentarily as percussionist (Fig. 9).
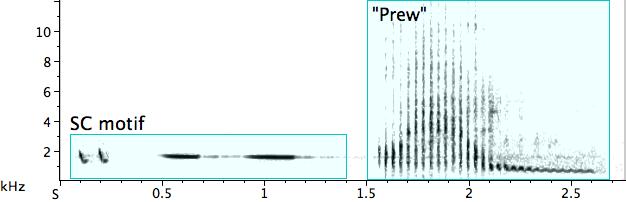
Fig. 9. Exemplar of the “short-long” motif, followed by an aggressive “prew” rattle.
Aside from the “prew” rattle, the “8-2-2” species call, and the “reek” alarm call, few mnemonics are in use for pied butcherbirds, likely because their song phrases lack stereotypy. Nevertheless, several have been posited: "weedle-ordle-ah-dit-dee" was applied to one phrase a recordist heard[xv], while “quo-orrr” and “chu-eep” (Serventy and Whittell 1976: 450) were pressed into service for certain notes. One final example, a veritable “period piece,” speaks as much about the writer as the birds:
His notes suggest the vastness of the Australian bush and continent. At dawn his clear cornet-like notes ring out far above the great chorus of bird-song. They are various, and some impossible of translation, but his dawn notes resemble the following:--‘Toll-de-lol-fãh’ (the last note long drawn out and of liquid sweetness); then twice and quickly repeated in a lower key—‘You chatterbox;’ then in a higher key and with very full, rounded notes, and twice repeated—‘Sweet after forty.’ So charmed was I with the song and appearance of these birds that I determined to secure one to take home with me. Through the kind offices of Mr. Lee Steere, I eventually managed to get one from one of the station hands, and my captive has furnished me with many opportunities of study (Milligan 1905: 154-155).
4. SONG
The purpose here is to present a sampler of analyses of songs and singing patterns observed in various contexts. An exhaustive account is not offered, and many omissions were necessary for reasons of space. Following Sloboda’s lead (1985: 9), my personal knowledge, experiences, and intuitions as a practicing musician have augmented and complemented the findings.
Rhythm: Rhythmic effects found in human music are often also present in pied butcherbird song, including accelerando, ritardando, rubato, syncopation, triplets and other tuplets, anacrusis, and additive and divisive rhythms. A number of durational values are present. The sense of a regular tempo, from adagio to allegro, and a rhythmic vitality are usually in evidence, and although to my metrical judgment there is a strong kinaesthetic component, the songs are not metronomically “impoverished” (Lidov 1997: 20) or brittle.
Bird songs are rhythmic enough, but they are seldom “measured” in the way that western music is measured. Music is regarded as measured when there is throughout a steady recurrence of rhythmic stress or accent so that the music is divided into regular measures or bars over a longish period. Very little bird utterance conforms to that description (Thorpe and Lade 1961: 231).
“Melody does not of necessity depend upon rhythm; like the songs of many birds, it can exist without recurrent, strictly gauged patterns of organization” (Sachs 1962: 111). In pied butcherbird song, patterns of organization regularly present themselves, but they are rarely strict.
We also find in the pied butcherbird a toolkit of expressions, articulations, and strategies familiar in the human animal’s music. While space does not permit a full listing, the following repertoire of musical procedures for varying sound is noted. In the absence of detailed experimental studies, inferring function is problematic. Martinelli argues that since singing, like speaking, is a communicative strategy, “…even aesthetics is a function in all respects. Not only is aesthetics not in competition with the other functions, but it often accompanies and enhances them” (2002: 12).
Canon. Countersinging, where neighbouring birds sing the same or similar songs along territory boundaries, can be of two sorts: “type-matching” involves the same phrase, while “repertoire-matching” draws on any song from the common repertoire (Beecher et al 2000: 21) The timing can be concurrent, alternating, or overlapping. When type-matching is in place, even if the entrances seem haphazard, a clear canonic treatment is established. (The result is a “performance” canon, not a notated one, reminiscent of Thelonius Monk and Charlie Rouse breaking into an improvised canon on a chorus of “Bemsha Swing.”[xvi]) While the function of countersinging could be a vocal contest (a “cutting session” in jazz parlance), a sure ploy for getting attention (playing on a rest beat), or a competitive advertisement (a commercial jingle) (Horn and Falls 1988: 337), the effect is a highly musical one. Anthropomorphism does not allow one to speculate on the possibility that creating a musical experience is even part of the function of countersinging, but almost anyone hearing it would dare to think so. Are pied butcherbird phrases formed with the instinct that they must work together when more than one singer in the avian song community is in earshot?
Crescendo/decrescendo. From my campground in the Central Australian outback, I heard a bird in the distance and decided to investigate the following day. At 3:45 a.m. under a full moon, I walked to where I thought it might sing and turned on my recorder. Five minutes later I heard birds quietly “warming up” nearby, and after another five minutes a bird flew into position in the tree above me and began to sing. I had stumbled on the box seat. The point of departure was the truncated version of a phrase delivered in hushed tones. The material was repeated, recombined, and elaborated for two hours. Mid-way through, the signal was so powerful it began to distort; I re-directed the microphone, but still the signal was fortissimo. By the end, the bird had brought his song phrases back to the original pianissimo, and the elaborated material was again presented in abridged form (Fig. 10).
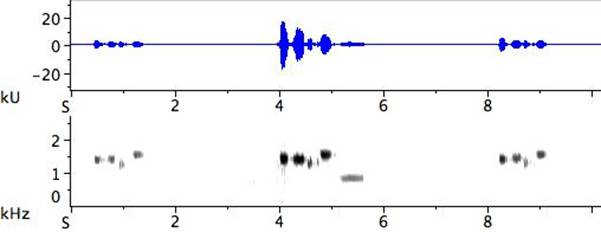
Fig. 10. Exemplar of crescendo/decrescendo: the first, a middling, and the final phrase from a two-hour pre-dawn song. The waveform on top shows the change in volume in more detail than the greyscale of the bottom sonogram.
Dawn Chorus. “Jazz is like bananas ¯ it must be consumed on the spot,” Sartre recommended (1948: 48). The active space of the dawn chorus is the ultimate surround-sound. Feathered choirs compete intensively for the available broadcasting space and time. The quality of the experience—the theatricity of this sonic story—is not entirely told by notation or sonograms. Space is an essential aspect in music (Brant 1967: 223-242). Vella speaks of aural depths of field—the foreground, middleground, background (2000: 132) and the shifting perception as sounds unfold. The acoustic tasks facing a bird in the dawn chorus are considerable. In the midst of competing “biophonies (voices of living things) and geophonies (non-creature sounds, e.g. thunder, rain, and wind)” (Krause 2002: xii), a singing bird must also assess other singers—their direction, distance, species, and identity—and who’s tuning in? It could be a predator. Why, then, risk expending an extravagant amount of energy (Smith 1991: 241) on song?
Fanfare.
“A flourish of trumpets…”(Tarr 2008). With cornet-like tone and bold, swift delivery, numerous pied butcherbird phrases aptly fit this metaphor. In Fig. 11, some portamenti were removed in order to highlight intervallic relationships—frequent, often rising major thirds and perfect fourths, fifths, and octaves.

Fig. 11. Exemplars of fanfares from seven different pied butcherbirds.
Iconic themes. Occasionally pied butcherbird themes and those of humans converge. Hutchinson reports hearing a phrase recalling the opening bars of Beethoven’s Fifth Symphony (Slater 1983: 278). I have encountered melodic inventions that correspond to Mendelssohn’s “Wedding March,” Kurt Weill’s “Mack the Knife,” and Miles Davis’s “Freddie Freeloader” (see Fig. 8). Although an iconic human theme can catch the ear, encountering an equally compelling and previously unknown theme provides greater satisfaction.
Melisma. Although they can be vocal gymnasts with a flourish here and an arabesque there, pied butcherbirds are not prone to extreme elaboration and appoggiatura. One from this pair is a notable exception, and the experience left a vivid impression with the recordist. This notation (Fig. 12) was accomplished both by listening at half-speed and by taking measurements in the sonogram window, particularly the latter.
“The Singing Lesson”: two birds perched side by side on a banksias singing a solo song; one sings the fully decorated version of the local dawn solo, the second a more sketchy outline version. A beautiful study of what structure, tuning and melodic decoration might mean to pied butcherbirds.[xvii]

Fig. 12. “The Singing Lesson,” where bird #1 puts forth a few basic notes (lower staff) while bird #2 unravels a melismatic cadenza (upper staff). (QR=quasi-rattle.)
Ostinato. Fig 13: In this dawn chorus a grey shrike-thrush (GST) Collurincia harmonica predominates, while the pied butcherbird (PBB) holds to a two-note ostinato. The harmonic implications make for a comfortable pairing. In another instance (not shown), the pied butcherbird inserts the only clearly tonal element into the acoustic fabric as it reiterates a two-note motif in an otherwise high-frequency shimmering of cicadas and “noisy” birdcalls.
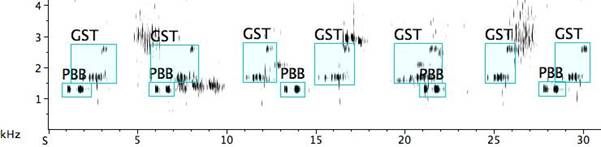

Fig. 13. Sonogram above of pied butcherbird ostinato whilst a grey shrike-thrush rings out its phrase, and below music notation of the first 10sec. (Other birds are also singing.)
Phrase endings.
The end of the phrase is usually differentiated rhythmically to provide punctuation. Phrase endings may be marked by a combination of distinguishing features, such as rhythmic reduction, melodic relaxation through a drop in pitch, the use of smaller intervals and fewer notes, or by any other suitable differentiation (Schoenberg 1967: 3).
Schoenberg’s suggestions are regular occurrences in pied butcherbird song (Fig. 14).

Fig. 14. Exemplars of phrase endings “á la Schoenberg”: 1. ¯ 3. rhythm reduction at measure’s end; 4. ¯ 6. a drop in the final pitch; 7. ¯ 9. smaller final intervals; 10. extended repetition of the final note; 11. a swift two octave rise with a half-step resolution deflected by the octave; 12. structural accent in the outline of a G dominant seven chord. (R=rattle.)
Scales. Scalar passages are not common. When scales arise (Fig. 15 and 16), they often consist of a combination of microtones, half and whole steps, and minor thirds; some notes can be repeated before the bird moves on. Other times, a “scale” is merely an ascending or descending rattle. These rare scales fail to suggest a sense of tonality or a tonic. No attempt has been made to piece together scales after the fact due to the prevalence of portamento.

Fig. 15. Exemplar of scalar motion in two different pied butcherbird songs.

Fig. 16. Exemplar of scalar motion from a pied butcherbird duo.
Sotto voce. The quiet, inward rendering of “whispering song” (Lister 1953: 142), also called whisper song, recording song, or quiet song, is most common during late mornings and afternoons. The material can be that of primary song (Sedgwick 1947: 377) or mimicry (Chisholm 1947: 25). Armstrong identifies this as the quietest form of song and speculates that it may have a variety of functions (1953: 311). One wonders why a bird would sing so quietly. With no apparent advertising function, is this subdued song merely a rehearsal (étude)? Could it be habit? Or could a bird simply like to sing—is it a self-rewarding activity (Morris 1962: 145)?[xviii]
Shape and balance. Hall-Craggs, a trained musician, begins the discussion section in her classic study of blackbird song with the apology that her sole qualification in the study of birdsong consists of “being trained to listen to detail” (1962: 294). She goes on to speak of the “shapeliness of melodic line and general formal orderliness” (ibid.: 296). She could have been speaking of Fig. 17. Songs that match the pied butcherbird’s perceptual world regularly match mine.

Fig. 17. Exemplar of shape and balance in four consecutive phrases of pied butcherbird song.

Fig. 18. Exemplar of shape and balance in four consecutive phrases of pied butcherbird song, with rests inserted between phrases to show approximate delivery.
Transposition. Birds’ hearing is acute in both low- and high-frequency ranges, their acoustic acuity and temporal discrimination is similar to human animals, and they are able to discriminate the songs of other species, even at the level of individuals, with precision (Greenewalt 1968: 138; Dooling 1982: 95-130; Dooling 1989: 423-444). European starlings possess absolute pitch and show a preference for learning pitch patterns on that basis (Hulse and Page 1988), while in the case of black-capped chickadees a melody can be retained with a change of key (Weisman et al 1990).
Do pied butcherbirds possess absolute pitch? By all accounts, yes. One bird was recorded almost daily over a two-and-a-half-week period in 2002, producing five hours of recordings. Two phrases with notes of almost constant frequency were chosen for every take, and the frequency at the beginning and ending of a song and at several mid-way points was measured. Results indicated a virtually imperceptible variation among renditions of the same phrase by the same individual (other measurements on different individuals produced similar results). Variations in recording technique, environmental conditions, and the position of the subject would be expected to introduce more variation than exists among the phrases as delivered.
Do pied butcherbirds transpose phrases (indicating relative pitch)? Yes. The numerous cases of the transposed species call are the clearest example of this; other examples were also noted.
Do pied butcherbirds possess octave generalisation? Do they perceive octaves as same or similar phenomena? Octave leaps abound in their song, both in the song of one bird and at the hand-off point in a duet. In another case, a pied butcherbird was singing in the presence of a more powerful signal from a magpie-lark Grallina cyanoleuca; the pied butcherbird mimicked the magpie-lark, transposing his phrase down an octave (possibly to a better range). It is tempting to speculate that stimulus equivalence for an octave could exist.
Timbre is an important but sometimes overlooked feature in vocal communication in birds (Williams, Cynx, and Nottebohm 1989: 379). Rhoades regretted the lack of a system of notation for timbre (1963: 200). For descriptive purposes, the voiceprint in a sonogram now fills the void, and in it we have a potent visual tool. The concept of Klangfarbenmelodie moves toward an equality of pitch and timbre, suggesting that a melody might be formed and perceived through timbral transformation of a single pitch. We find such instances in pied butcherbird song. In Fig. 19, Example 1, the phrase begins with a quasi-rattle, followed by a pure tone, and finishing with a proper rattle that moves back to a quasi-rattle, all on a single pitch. The second example finds a bird replacing its beginning sustained tone with extreme portamento signals recalling the vintage VCS-3 synthesizer[xix]. These much shorter notes begin quietly and crescendo slightly, but are in stark dynamic disparity to the powerful final octave and a half jump down to the beginning pitch. Although not limited to a single pitch, Example 3 offers extreme timbre differentiation. Parameters exhibiting strong contrast include note durations, dynamics, and texture—alternating pure notes and noisy ones, smoothness and roughness (Malloch 2004: 54-56). Intensification via powerful attacks and steep but rapid ascending and descending portamento is in evidence, and again quasi-electronic signals come to mind. Finally, each of the fourth bird’s virtuosic song phrases displays clear, bold quality differentiation, at times imparting a feeling of impulsiveness. The palette is remarkably wide-ranging for one bird, with strong fluctuations in multiple parameters.
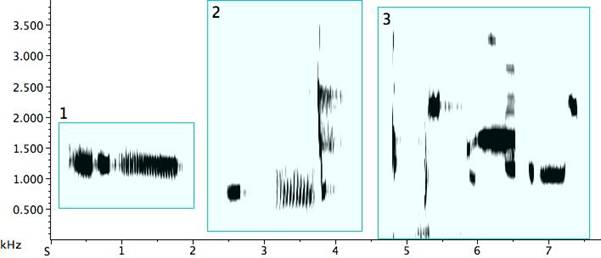
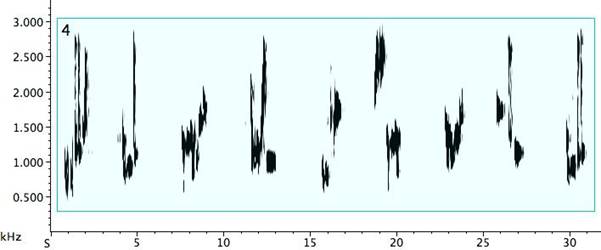
Fig. 19. Four exemplars of timbre contrast in pied butcherbird song.
Ventriloquism. The ability to make sounds appear to come from somewhere other than the emitter occurs in the alarm calls of some birds as a defense mechanism (Marler 1955: 7). Ventriloquism is a known occurrence in the dawn song of the pied butcherbird (Keast 1944: 185). Its function in birdsong is not known. Imagine hearing a number of individuals quietly participating in the dawn chorus, echoing similar themes, then perhaps all shifting to a new theme; as you close in (you think), like the end of a rainbow, no bird can be found. The sound has no address. Normally not considered a musical activity, ventriloquism nevertheless possesses musical connotations and precedents.[xx]
Repetition and variation. The hook, the Ohrwurm (earworm), the brainworm[xxi]—whatever the terminology, these catchy bits that stick in our head and endlessly repeat—do they have a counterpart in the avian world? Are pied butcherbird song phrases designed to have such an effect? Fig. 20 illustrates a simple, repetitive theme that commands attention and is remembered (some portamenti were removed for ease of visual inspection).

Fig. 20. A catchy “hook” from a pied butcherbird.
Pied butcherbird songs are regularly constructed by rearranging and recombining song units, creating tension, contrast, and balance. Do short or long strings of master songs appear as organisational or memory strategies? When large amounts of vocal material are subjected to distributional analysis and inspected, no predictable phrase ordering can be detected. Very little research into “long” birdsongs has been undertaken. For the pied butcherbird, a “long” song would be a minimum thirty minutes’ duration up to three hours or more; the recording would be made without interruption and be composed of the entire vocalisation delivered by one bird. Continuing fieldwork will involve recording as many coherent long song cycles as possible. Capturing the entire performance involves the serendipity and skill of being in place and commencing the recording before the bird begins to sing.
The following questions are of interest: What are the underlying rules that direct and structure the song? Are there similar principles of organisation and development between pre-dawn and day song? Are improvisation and invention applied to certain phrases or motifs while other segments remain unchanged? What features or segments serve as personal identification markers? What are the most stable parameters/most fluctuating parameters? What kind of potential permutations are never observed? What length of recording is necessary to hear all the phrases in a repertoire?
Repertoire. Song delivery, repertoire sizes, and organization of repertoires were examined. A bird with a repertoire of four or more distinct phrases is termed a “versatile” singer (versus “non-versatile”); a bird that sings more than 50% of the time during a normal performance period is categorised as a “continuous” singer (versus “discontinuous”) (Hartshorne 1956: 176-177). Pied butcherbirds would be considered versatile, discontinuous singers. Determining the total number of different song components in a repertoire is both difficult and arbitrary (Kroodsma 1977: 998). In the course of one song performance, a pied butcherbird often works with 4-6 phrases; by the time material has been deleted, recombined, and otherwise reworked, especially if the recording is over thirty minutes, one is hard-pressed to count what is a permutation from what is a new phrase. It would seem that their acceptance and rejection of ideas (as well as transformation) are consistent with the ability to compose (Hold 1971: 114), and not merely the “manifestation of their behavioural flexibility” (Baptista and Trail 1992: 245).
Most phrases are 1¯2sec. in duration. Sometimes only one note is sung (around .2sec.) A phrase of 4.5sec. is the longest coherent phrase in my library (with no pause and no possibility of another bird also singing). The normal rate of delivery when vocalising maximally could be 8-16 phrases/minute; a high rate of delivery would be 17-22 or more. (This rate is dependent on the length of the recording. Over a two-hour period, for example, this rate of delivery would fall. Recordists often only send me their “best” material, perhaps covering three to ten minutes of peak activity. A more typical example would be a pre-dawn song of 110 minutes that saw 556 phrases delivered, or 5/minute.) The inter-phrase interval could be expected to be as long as the phrase it follows, often twice as long or more. Does this gap between phrases simply mark a switch in function from singing to listening, or is it part of the phrase? Hartshorne included these silent periods between phrases as part of the duration of the phrase, believing that feathered songsters seek a balance in performance between sound and silence (1956: 177). Sing-then-listen is akin to the human improviser’s art.
Stage presence.
It is late afternoon on the border of Western Australia and the Northern Territory. While stopped, my partner photographs a pied butcherbird on an overhead wire with quite a repertoire, one who tilts back his head, opens his bill, and puffs out in song, a born performer who turns on for the camera. No other birds are apparent. I grab my paper to notate his call as he delivers a dynamic singing performance. He had “it.” (Taylor 2007: 51).
Duetting, antiphonal song, and female song. Birdsong is often presented as a contest between rival males or between the skilled salesmanship of a male versus the equally well-developed sales resistance of a female (Williams 1966: 184). This functional definition glosses over the fact that females sing much more than is usually recognized (Smith 1991: 248). Females may sing solo or in duets (Langmore 1998) and are known to sing to attract mates (Langmore 1998; 2000). Their repertoire may be as large or larger than males (Brown and Farabaugh 1991: 270-271).
Just over 200 of the world’s 9,000 or so bird species are known to duet (Hall 2002-2003: 53). The function of duetting, like that of female song, remains puzzling, although various hypotheses have been explored, e.g. pair bond maintenance, mutual stimulation, contact, cooperative territorial defense, and mate guarding (Thorpe 1972; 1973: 73; Grafe et al 2004: 181; Hall 2004; Rogers et al 2007). At times two or more pied butcherbirds may exchange loosely overlapping bouts of song phrases and calls. Duetting occurs when two individuals, assumed to be a mated pair or potential mates, coordinate their vocalisations in a synchronous or alternating pattern (Mulder et al 2003: 25). Antiphonal duetting implies considerable speed and precision in the alternating of motifs (Power 1966: 314). Such precision calls to mind the dovetailing of the medieval hocket.
Female song plays a key role in duets and antiphonal song for pied butcherbirds; unfortunately, since the sexes are indistinguishable, the female contribution can only be speculated on. Unison duets are rare, although in trios unison motifs are not uncommon. The timing of antiphonal duets is normally such that without visual contact, the contributions do not parse easily and must be noted by the recordist in the field. When singing, pied butcherbirds often alternate a standard upright posture with raising the bill high, and then sinking it on the breast, which assists in part identification.
The distinctive duet of a mated pair of Pied Butcher-Birds is among the sweetest music of bush birds. While performing, the pair courtesy to each other, and at the same time are answering other songful pairs, so that the woodland resounds, as it were, with a round of music (Campbell and Barnard 1917: 38).
Further studies could shed light on whether duets make use of entirely separate material or whether motifs from solo songs are converted into duets.
The joint performance in Fig. 21 involves rapid exchanges. Of interest is the transposition of the second motif D ¯ C by an octave. The final motif is also held in common by both birds and resembles the first: the D ¯ G# motion is approached chromatically in the first case and by alternating thirds in the latter two. On the morning of this recording, this same pair sang three other duets, all containing different material.

Fig. 21. Antiphonal song from two birds.
Mimicry. Composers borrow from one another, and even from their own previous works; jazz improvisers quote themes from both within and outside jazz; pied butcherbirds appropriate from conspecifics, other bird species, and unexpected sources. Mimicry, or imitation, is the ability to reproduce, to a varying degree, sounds other than those of the species in question (Lemaire 1975: 95), including environmental sounds. Its function is poorly understood, and no single explanation appears to suffice (Chisholm 1946; Marshall 1950; Baylis 1982; Kroodsma 2004: 128-131). Therefore, the definition given is a descriptive rather than a functional one.
The literature enumerates 27 bird species apparently taken up with relish by the pied butcherbird, as well as the barking of a dog, the bleating of a lamb, a person whistling (Higgins, Peter, and Cowling 2006: 522), and the neighing of a horse (Lord 1945: 119). The recordings in my library find the pied butcherbird mimicking nine bird species from this list. The imitations are short, of about 1-2sec. duration, making the identification challenging or sometimes impossible. With the expert assistance of several ornithologists[xxii], my research adds 12 bird species to this list, for a total of 39 species, plus six probable new bird species and a frog, as well as credible mimicry of a human voice, a telephone bell, and the meow of a cat. (Approximately 25% of the apparent mimicry remains unidentified.) Such aural and vocal achievement is impressive. Most remarkable, one bird mimics an auditory icon: the signal of a reversing truck is matched in frequency, duration, and timbre, and incorporated into dawn song. (The truck was recorded later that morning at a construction site adjacent to the bird’s songpost.) This indicates that they hear sound as we do. Fig. 22 pairs bird/truck and bird/telephone.
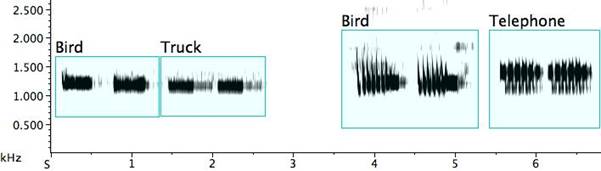
Fig. 22. Exemplar of mimicry: a pied butcherbird and the signal of a reversing truck in the bird’s territory; exemplar of possible mimicry: a pied butcherbird and the signal of a ringing telephone audible from outdoors in its territory.
While sometimes one motif from an alien species might be absorbed into a pied butcherbird phrase, other times they string together a postmodern pastiche of imitations in a mimicry cycle. Supposed norms are violated: their vocal range extends in both directions (the normal singing range encompasses at least a two-octave span, from 698 (F5) to 2794 (F7) Hz; the mimicry singing range covers a three-and-a-half octave span, from 506 (B4) to 5706 (F8) Hz), and there is often no inter-phrase interval (the rate of delivery goes up to 30/minute, and in one example a bird delivers at 34/minute); the bird might sing nearly nonstop for fifteen minutes or more. The mimicry cycle is usually delivered quietly. Other assumed constraints in singing ability or singing affinity are shown to be plastic. For example, simple up-and-down warbles might be exaggerated (Fig. 23). The full-on effect is one of a DJ cut-and-paste session. Why would a bird transgress its species-specific sounds with these acoustic abstractions? One assumes this degree of complexity and elaboration would be well beyond what is necessary for survival and reproduction, suggesting yet again the presence of aesthetic appreciation (Hartshorne 1973: 56; Jellis 1977: 204).
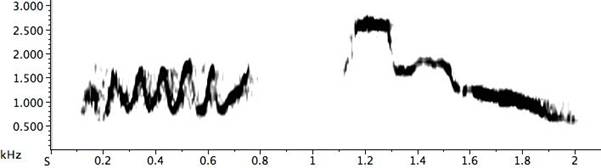
Fig. 23. Exemplars of extreme warbling notes from a pied butcherbird mimicry cycle.
Conclusion.
Part of the essence of being a pied butcherbird is song. They have a keen interest in sound patterns and take pains to develop skill in their production. Although it can include mimicry of a sonic event from a human source, their song is not sullied by commerce. It has survived contamination by the culture industry. As we hurtle towards a reductive global culture, their vocalisations enjoy authenticity, “the cultural ideal” (Nettl 1983: 318). Their songs are sonic heirlooms that look back to human pre-history and look forward: their melodic inventions are dynamic, in a state of flux and constant repositioning. Variations are found at all levels of organisation. Many components from their rich and nuanced repertoire are subject to recasting, some via elaborate strategies that seemingly overreach biological necessity. This panoply of recombining, varying, and inventing mechanisms causes me to believe that aesthetic statements are being delivered and that the birds appreciate this in their way. “If invention can reveal itself as being as important in the individuals of other species as in man, it is time to get rid of the image (three centuries old) of animal-machines” (Mâche 1983/1997: 157). Pied butcherbirds—on the continuum from automatons to science experiments to fellow musicians—where will we place them?
Endnotes
- [i] “It is best to keep an open mind about the possibility of consciousness in all animals that exhibit versatile behavior or communicate in ways that suggest they may be expressing thoughts or feelings” (Griffin 1992: 4). Since as yet we have no “general theory” of human cognition, it is understandable that scientists who study animal behaviour avoid this subject. Little is known, but it would seem anthropomorphic not to entertain the possibility of animal consciousness.
- [ii] Rogers and Kaplan (2000: 40-42) explore the basic concept of territorial songs, while Martinelli (2002: 25-28) examines the theory of Umwelt, moving from the physical environment to the phenomenological one.
- [iii] “It would be interesting to ask the same questions of human song as are asked of bird song: Why do humans engage in song? Pair bond maintenance? Mutual stimulation? Contact? Cooperative territorial defense? Mate guarding? Does the probability that such goals are present indicate that humans cannot be considered to demonstrate a proper musical aesthetic?” Personal communication dated 21 December 2007 from Bob Ostertag, Associate Professor of Technocultural Studies, University of California, Davis.
- [iv] Personal communication dated 21 June 1989 from Olivier Messiaen to Australian ornithologist and lyrebird expert Syd Curtis.
- [v] This difference in approach between biologists and musicologists is reminiscent of the tug-of-war in ethnomusicology between anthropologists and musicologists (Merriam 1969).
- [vi] The software Amadeus is available at http://www.hairersoft.com/ (accessed 10 April 2008).
- [vii] The software Raven is specifically designed for the analysis of animal acoustics and is available at http://www.birds.cornell.edu/brp/raven/Raven.html (accessed 10 April 2008).
- [viii] Burton encountered similar difficulties in notating Native American songs with portamento, bemoaning the “vagueness” (1909: 22).
- [ix] In a chapter contemplating the influence of birds on the origins of music, Pont (1988) describes a preliminary statistical study of melodic contour preferences and proposes the comparison of human and avian pitch profiles as tools for the investigation of possible regional co-variation.
- [x] Personal communication dated 10 August 2006 from François-Bernard Mâche.
- [xi] The software Finale is available at http://www.makemusic.com/ (accessed 10 April 2008).
- [xii] A pied butcherbird approached me at an outdoor café where I was lunching on fish and chips. I handed over a chip while I grabbed the recorder; not a vegetarian, s/he waited until I offered the preferable fish course. Once the bird had fed, it took what it could not finish and impaled it on a fork in a tree, and thus the bird’s name; like a butcher, it stores surplus food.
- [xiii] Personal communication dated 2 July 2005 from composer and recordist David Lumsdaine.
- [xiv] Octave designations are based on those used by the software Amadeus II, which conform to the Acoustical Society of America System.
- [xv] Personal communication dated 12 March 2007 from wildlife recordist and ornithologist Vicki Powys.
- [xvi] From Thelonius Monk: The Complete Riverside Recordings, 1986, Riverside RCD-022-2, disc 15, track 8.
- [xvii] Personal communication dated 2 July 2005 from David Lumsdaine.
- [xviii] Humans are known to sing to themselves when they are alone.
- [xix] The VCS-3 is a portable synthesizer designed in 1969. The instrument is documented at http://emfinstitute.emf.org/exhibits/vcs3.html (accessed 10 April 2008).
- [xx] The offstage choruses in Verdi’s operas Il Corsaro and Il Trovatore come to mind, as does the remarkable soprano Edith Helena who mimicked the violin with her voice while appearing onstage playing a stringless violin in pantomime to “Intermezzo from Cavalleria Rusticana” (“The London Music Halls ¯ The Empire.” The Era, 23 January 1904: 23).
- [xxi] Levitin (2006: 151) and Sacks (2007: 41-48) discuss this phenomenon in more detail.
- [xxii] Australian ornithologists Syd Curtis, Gayle Johnson, Vicki Powys, Carol Probets, and Eric Vanderduys assisted in mimicry identification.
Bibliography
- Adret, Patrice. 2004. “In Search of the Song Template.” In Behavioral Neurobiology of Birdsong, eds. H. Philip Zeigler and Peter Marler, 303-324. New York City: Annals of the New York Academy of Sciences.
- Armstrong, Edward A. 1953. "Secondary song." British Birds 46: 311.
- Armstrong, Edward A. 1973. A Study of Bird Song. New York: Dover Publications.
- Baptista, Luis and Keister, R. A. 2005. "Why birdsong is sometimes like music." Perspectives in Biology and Medicine 48(3): 426-443.
- Baptista, Luis F. 1975. "Song dialects and demes in sedentary populations of the white-crowned sparrow (Zonotrichi leucophrys nuttalli)." University of California Publications in Zoology 105: 1-52.
- Baptista, Luis F. and Gaunt, Sandra L. 1997. “Social interaction and vocal development in birds.” In Social Influences on Vocal Development, eds. Charles T. Snowdon and Martine Hausberger, 23-40. Cambridge, U.K.: Cambridge University Press.
- Bartók, Béla and Lord, Albert B. 1951. Serbo-Croatian Folk Songs. New York: Columbia University Press.
- Baptista, Luis F. and Trail, Pepper W. 1992. "The role of song in the evolution of passerine diversity." Systematic Biology 41(2): 242-247.
- Baylis, Jeffrey R. 1982. “Avian Vocal Mimicry: Its Function and Evolution.” In Acoustic Communication in Birds: Song Learning and Its Consequences, Vol. 2, eds. Donald E. Kroodsma, Edward H. Miller, and Henri Ouellet, 51-83. New York: Academic Press
- Beecher, Michael D. et al. 2000. "Territory tenure in song sparrows is related to song sharing with neighbours, but not to repertoire size." Animal Behaviour 59(1): 29-37.
- Blacking, John. 1995. How Musical Is Man?. Seattle: University of Washington Press.
- Bowman, Wayne D. 1998. Philosophical Perspectives on Music. New York: Oxford University Press.
- Boyden, David D. and Stowell, Robin: “Glissando.” In Grove Music Online, ed. L. Macy, <http://www.grovemusic.com> [accessed 17 July 2007].
- Bradbury, Jack W. and Vehrencamp, Sandra L. 1998. Principles of Animal Communication. Sunderland, Massachusetts: Sinauer Associates.
- Brant, Henry. 1967. “Space as an Essential Aspect of Musical Composition.” In Contemporary Composers on Contemporary Music, eds. Eliot Schwartz and Barney Childs, 221-242. New York: Holt, Rinehart and Winston.
- Broughton, W.B. 1963. “Method in Bio-Acoustic Terminology.” In Acoustic Behaviour of Animals, ed. R.-G. Busnel, 3-24. Amsterdam: Elsevier.
- Brown, Eleanor D. and Farabaugh, Susan M. 1991. "Song sharing in a group-living songbird, the Australian magpie, Gymnorhina tibicen. Part III. Sex specificity and individual specificity of vocal parts in communal chorus and duet songs." Behaviour 118(3-4): 244-274.
- Burton, Frederick R. 1909. American Primitive Music. New York: Moffat, Yard and Company.
- Campbell, A. J. and Barnard, H. G. 1917. "Birds of the Rockingham Bay district, North Queensland." Emu 17(1): 2-38.
- Catchpole, C. K. and Slater, P. J. B. 1995. Bird Song: Biological Themes and Variations. Cambridge, U.K.: Cambridge University Press.
- Chisholm, A. H. 1946. Nature's Linguists: A Study of the Riddle of Vocal Mimicry. Melbourne: Brown, Prior, Anderson.
- Chisholm, A. H. 1947. "More about vocal mimics." Victorian Naturalist 64: 25-26.
- Cook, Nicholas. 1998. Music: A Very Short Introduction. Oxford: Oxford University Press.
- Covell, Roger. 1967. Australia's Music. Melbourne: Sun Books.
- Craig, Wallace. 1943. The Song of the Wood Pewee Myiochanes virens linnaeus: A Study of Bird Music. Albany, New York: The University of the State of New York.
- Cross, Ian. 2003. “Music as a Biocultural Phenomenon.” In The Neurosciences and Music, ed. Guiliano Avanzini et al, 106-111. New York: The New York Academy of Sciences.
- Dooling, Robert J. 1982. “Auditory Perception in Birds.” In Acoustic Communication in Birds: Song Learning and Its Consequences, Vol. 2, eds. Donald E. Kroodsma, Edward H. Miller, and Henri Ouellet, 95-130. New York: Academic Press.
- Dooling, Robert J. 1989. “Perception of Complex, Species-Specific Vocalizations by Birds and Humans.” In The Comparative Psychology of Audition: Perceiving Complex Sounds, eds. Robert J. Dooling and Stewart H. Hulse, 423-444. Hillsdale: Lawrence Erlbaum Associates.
- Doupe, Allison J. and Kuhl, Patricia K. 1999. "Birdsong and human speech: common themes and mechanisms." Annual Review of Neuroscience 22: 567-631.
- Feynmann, Richard P. 1999. The Pleasure of Finding Things Out. London: Allen Lane.
- Fitch, W. Tecumseh. 2006. "The biology and evolution of music: a comparative perspective." Cognition 100(1): 173-215.
- Fletcher, Neville H. 1992. Acoustic Systems in Biology. New York: Oxford University Press.
- Fletcher, Neville H. 2006. "Birdsong science." Australian Science 27(9): 35-37.
- Gaunt, Abbot S. 1983. "An hypothesis concerning the relationship of syringeal structure to vocal abilities." The Auk 100: 853-862.
- Goller, Franz and Larsen, Ole N. 1997. "A new mechanism of sound generation in songbirds." Proceedings of the National Academy of Sciences, USA 94(26): 14878-14791.
- Goller, Franz and Larsen, Ole N. 2002. "New perspectives on mechanisms of sound generation in songbirds." Journal of Comparative Physiology A 188: 841-850.
- Gourlay, K. A. 1978. "Towards a reassessment of the ethnomusicologist's role in research." Ethnomusicology 22(1): 1-35.
- Grafe, T. Ulmar; Bitz, Johannes H.; and Wink, Michael. 2004. "Song repertoire and duetting behaviour of the tropical boubou, Laniarius aethiopicus." Animal Behaviour 68(1): 181-191.
- Greenewalt, C. H. 1968. Bird Song: Acoustics and Physiology. City of Washington: Smithsonian Institution Press.
- Griffin, Donald R. 1992. Animal Minds. Chicago: The University of Chicago Press.
- Hall, Michelle L. 2004. "A review of hypotheses for the functions of avian duetting." Behavioral Ecology and Sociobiology 55(5): 415-430.
- Hall, Michelle. 2002-2003. "Duetting larks." Nature Australia 27(7): 52-59.
- Hall-Craggs, Joan. 1961. “The Aesthetic Content of Bird Song,” In Bird-song: The Biology of Vocal Communication and Expression in Birds, ed. W. H. Thorpe, 367-381. Cambridge, U.K.: Cambridge at the University Press.
- Hall-Craggs, Joan. 1962. "The development of song in the blackbird." Ibis 104(3): 277-300.
- Haraway, Donna J. 1991. Simians, Cyborgs, and Women: The Reinvention of Nature. London: Free Association Books.
- Harrison, Frank. 1977. "Universals in music: towards a methodology of comparative research." The World of Music: Quarterly Journal of the International Institute for Comparative Music Studies and Documentation 19(1/2): 30-36.
- Hartshorne, Charles. 1953. "Musical values in Australian songbirds." Emu 53: 109-128.
- Hartshorne, Charles. 1956. "The monotony-threshold in singing birds." The Auk 73: 176-192.
- Hartshorne, Charles. 1958. "The relation of bird song to music." Ibis 100: 421-445.
- Hartshorne, Charles. 1973. Born to Sing: An Interpretation and World Survey of Bird Song. Bloomington, Indiana: Indiana University Press.
- Higgins, P. J.; Peter, J. M.; Cowling, S.J. (eds.). 2006. Handbook of Australian, New Zealand & Antarctic Birds Vol 7(A). Melbourne: Oxford University Press.
- Hold, Trevor. 1971. "Messiaen's birds." Music & Letters 52(2): 113-122.
- Horn, Andrew G. and Falls, J. Bruce. 1988. "Repertoires and countersinging in western meadowlarks (Sturnella neglecta)." Ethology 77: 337-343.
- Hulse, Stewart H. and Page, Suzanne C. 1988. "Toward a comparative psychology of music perception." Music Perception 5(4): 427-452.
- Hultsch, Henrike and Todt, Dietmar. 2004. “Learning to sing.” In Nature's Music: The Science of Birdsong, eds. Peter Marler and Hans Slabbekoorn, 80-107. San Diego: Elsevier Academic Press.
- Jellis, Rosemary. 1977. Bird Sounds and Their Meaning. Ithaca, New York: Cornell University Press.
- Johnson, Gayle. 2003. Vocalizations in the grey butcherbird Cracticus torquatus with emphasis on structure in male breeding song: implications for the function and evolution of song from a study of a Southern Hemisphere species. Australian School of Environmental Studies. Nathan, Queensland: Griffith University. Doctor of Philosophy.
- Jurisevic, M. A. and Sanderson, K. J. 1994. "Alarm vocalisations in Australian birds: convergent characteristics and phylogenetic differences." Emu 94: 69-77.
- Keast, J. A. 1944. "A winter list from the Tweed River district, NSW, with remarks on some nomadic species." Emu 43: 177-187.
- Kivy, Peter. 1990. Music Alone: Philosophical Reflections on the Purely Musical Experience. Ithaca: Cornell University Press.
- Kolinski, Mieczyslaw. 1967. "Recent trends in ethnomusicology." Ethnomusicology 11(1): 1-24.
- Krause, Bernie. 2002. Wild Soundscapes: Discovering the Voice of the Natural World. Berkeley: Wilderness Press.
- Kroodsma, Don. 2004. “The diversity and plasticity of birdsong.” In Nature's Music: The Science of Birdsong, eds. Peter Marler and Hans Slabbekoorn, 108-131. San Diego: Elsevier Academic Press.
- Kroodsma, Donald E. 1977. "Correlates of song organization among North American wrens." The American Naturalist 111(981): 995-1008.
- Langmore, Naomi E. 1998. "Functions of duet and solo songs of female birds." Trends in Ecology & Evolution 13(4): 136-140.
- Langmore, Naomi E. 2000. “Why Female Birds Sing.” In Animal Signals: Signalling and Signal Design in Animal Communication, eds. Y. Espmark, T. Amundsen, and G. Rosenqvist, 317-327. Trondheim, Norway: Tapir Academic Press.
- Lemaire, Françoise. 1975. "Dialectal variations in the imitative song of the marsh warbler (Acrocephalus palustris) in western and eastern Belgium." Le Gerfaut 65: 95-106.
- Levitin, Daniel J. 2006. This Is Your Brain on Music: The Science of a Human Obsession. New York, Dutton.
- Lidov, David. 1997. "Our time with the druids: what (and how) we can recuperate from our obsession with segmental hierarchies and other 'tree structures'." Contemporary Music Review 16(4): 1-28.
- List, G. 1974. "The reliability of transcription." Ethnomusicology 18(3): 353-377.
- List, G. 1979. "Ethnomusicology: a discipline defined". Ethnomusicology 23(1): 1-4.
- Lister, M. D. 1953. "Secondary song: a tentative classification." British Birds 46(8): 139-143.
- Lord, E. A. R. 1945. "Cracticus nigrogularis (black-throated butcher bird) as mimic." Queensland Naturalist XII(6): 118-119.
- Lynch, Alejandro et al. 1989. "A model of cultural evolution of chaffinch song derived with the meme concept." The American Naturalist 133(5): 634-653.
- Mâche, François-Bernard. 1983/1997. Music, Myth and Nature. Switzerland: Harwood Academic Publishers.
- Mâche, François-Bernard. 2000. “The Necessity of and Problems with a Universal Musicology.” In The Origins of Music, eds. Nils L. Wallin, Björn Merker, and Steven Brown, 473-479. Cambridge, Massachusetts: MIT Press.
- Malloch, Stephen. 2004. "An exploration of timbre analysis: the game of sound in two performances of Jeux vénitiens." Musicae Scientiae VIII(1): 53-81.
- Marler, Peter and Peters, Susan. 1982. "Structural changes in song ontogeny in the swamp sparrow Melospiza georgiana." The Auk 99: 446-458.
- Marler, Peter. 1955. "Characteristics of some animal calls." Nature 176(4470): 6-8.
- Marler, Peter. 1981. "Birdsong: the acquisition of a learned motor skill." Trends in Neurosciences 4: 88-94.
- Marler, Peter. 1994. “The instinct to learn.” In Language Acquisition: Core Readings, ed. P. Bloom, 591-617. Cambridge MA: The MIT Press.
- Marler, Peter. 1997. "Three models of song learning: evidence from behavior." Journal of Neurobiology 33: 501-516.
- Marler, Peter. 2004. “Bird Calls: Their Potential for Behavioral Neurobiology.” In Behavioral Neurobiology of Birdsong, eds. Philip H. Zeigler and Peter Marler, 31-44. New York City: Annals of the New York Academy of Sciences.
- Marshall, A. J. 1950. "The function of vocal mimicry in birds." Emu 50: 5-16.
- Martinelli, Dario. 2001. "Symptomatology of a semiotic research: methodologies and problems in zoomusicology." Sign Systems Studies 29(1): 1-12.
- Martinelli, Dario. 2002. How Musical Is a Whale? Towards a Theory of Zoömusicology. Hakapaino: International Semiotics Institute.
- McClary, Susan. 2000. Conventional Wisdom: The Content of Musical Form. Berkeley: University of California Press.
- Merriam, Alan P. 1963. "Purposes of ethnomusicology, an anthropological view." Ethnomusicology 7(3, Tenth Anniversary Issue): 206-213.
- Merriam, Alan P. 1963. "Purposes of ethnomusicology, an anthropological view." Ethnomusicology 7(3, Tenth Anniversary Issue): 206-213.
- Merriam, Alan P. 1964. The Anthropology of Music. Evanston, Illinois: Northwestern University Press.
- Merriam, Alan P. 1969. "Ethnomusicology revisited." Ethnomusicology 13(2): 213-229.
- Milligan, Alex W. 1905. "Notes on a trip to the Yandanooka district, Western Australia." Emu 4(4): 151-157.
- Morris, Desmond. 1962. The Biology of Art. London: Methuen & Co.
- Mulder, Raoul A., et al. 2003. "Alternate functions for duet and solo songs in magpie-larks, Grallina cyanoleuca." Australian Journal of Zoology 51: 25-30.
- Nettl, Bruno. 1983. The Study of Ethnomusicology: Twenty-nine Issues and Concepts. Urbana: University of Illinois Press.
- Nottebohm, Fernando. 1971. "Neural lateralization of vocal control in a passerine bird." The Journal of Experimental Zoology 177: 229-262.
- Nowicki, Stephen and Marler, Peter. 1988. "How do birds sing?" Music Perception 5(4): 391-426.
- Pont, Graham. 1988. “The Origin of Music: new light on an old hypothesis.” In Orígenes de la Música y Biomusicología: La Música Vista por la Filosofía y la Ciencia (Music Origins and Biomusicology: Music Seen by Philosophy and Science), ed. Julián Monge-Nájera, 29-35. San José, Costa Rica: Universidad Estatal a Distancia.
- Power, Dennis M. 1966. "Antiphonal duetting and evidence for auditory reaction time in the Orange-chinned Parakeet." The Auk 83: 314-319.
- Prum, Richard O. 1998. "Sexual selection and the evolution of mechanical sound production in manakins (Aves: Pipridae)." Animal Behaviour 55(4): 977-994.
- Rhoades, Willard. 1963. "Musicology and musical performance (Comments on Hood, "Musical significance")." Ethnomusicology 7(3, Tenth Anniversary Issue): 198-200
- Richman, Bruce. 1987. "Rhythm and melody in gelada vocal exchanges." Primates 28(2): 199-223.
- Rogers, Amy C.; Langmore, Naomi E.; and Mulder, Raoul A. 2007. "Function of pair duets in the eastern whipbird: cooperative defense or sexual conflict?" Behavioral Ecology 18(1): 182-188.
- Rogers, Lesley J. and Kaplan, Gisela. 2000. Songs, Roars, and Rituals: Communication in Birds, Mammals, and Other Animals. Cambridge: Massachusetts: Harvard University Press.
- Rothenberg, D. 2005. Why Birds Sing: A Journey into the Mystery of Bird Song. New York: Basic Books.
- Sachs, Curt. 1962. The Wellsprings of Music. New York: Da Capo Paperback.
- Sacks, Oliver. 2007. Musicophilia: Tales of Music and the Brain. New York, Alfred A. Knopf.
- Sartre, Jean-Paul. 1947. "I discovered jazz in America." Saturday Review of Literature 29 November 1947: 48-49.
- Schoenberg, Arnold. 1967. Fundamentals of Musical Composition. London: Faber and Faber Limited.
- Seashore, Carl E. 1938/1967. Psychology of Music. New York: Dover.
- Sedgwick, Eric H. 1947. "Northern Territory bird notes." Emu 46: 349-378.
- Serventy, D. L. and Whittell, H. M. 1976. Birds of Western Australia. Perth: University of Western Australia Press.
- Sessions, Roger. 1950/1974. The Musical Experience of Composer, Performer, Listener. Princeton: Princeton University Press.
- Shiovitz, Kenneth A. 1975. "The process of species-specific song recognition by the indigo bunting, Passerina cyanea, and its relationship to the organization of avian acoustical behavior." Behaviour 55: 128-179.
- Siegfried, T. 2006. A Beautiful Math: John Nash, Game Theory, and the Modern Quest for a Code of Nature. Washington, D.C.: Joseph Henry Press
- Slater, Peter. 1983. A Field Guide to Australian Birds. Adelaide: Rigby.
- Sloboda, John A. 1985. The Musical Mind: The Cognitive Psychology of Music. Oxford: Oxford University Press.
- Smith, W. John. 1991. "Singing is based on two markedly different kinds of signaling." Journal of Theoretical Biology 152: 241-253.
- Sotavalta, Olavi. 1956. "Analysis of the song patterns of two sprosser nightingales, Luscinia luscinia." Annals of the Finnish Zoological Society "Vanamo" 17(4): 1-31.
- Spector, David A. 1994. "Definition in biology: the case of 'bird song'." Journal of Theoretical Biology 168: 373-381.
- Stevens, Catherine. 2004. "Cross-cultural studies of musical pitch and time." Acoustical Science & Technology 26(6): 433-38.
- Suchoff, Benjamin. 1976. Béla Bartók Essays. Lincoln: University of Nebraska Press.
- Taylor, Hollis. 2005. “A call of the pied butcherbird.” AudioWings 8(2): 4-8.
- Taylor, Hollis. 2006. “Frequency measurements on a phrase of the pied butcherbird.” AudioWings 9(2): 15-18.
- Taylor, Hollis. 2007. Post Impressions: A Travel Book for Tragic Intellectuals. Portland, Oregon: Twisted Fiddle.
- Tarr, Edward H.: “Fanfare.” In Grove Music Online, ed. L. Macy, <http://www.grovemusic.com> [accessed 10 April 2008].
- Thompson, Nicholas S.; LeDoux, Kerry; and Moody, Kevin. 1994. "A system for describing bird song units." Bioacoustics 5: 267-279.
- Thorpe, W. H. 1958. "Further studies on the process of song learning in the chaffinch (Fringilla coelebs gengleri)." Nature 182: 554-557.
- Thorpe, W. H. 1972. Duetting and Antiphonal Song in Birds: Its Extent and Significance. Leiden: E. J. Brill.
- Thorpe, W. H. 1973. "Duet-singing birds." Scientific American 229(2): 70-79.
- Thorpe, W. H. and Lade, B. I. 1961. "The songs of some families of the passeriformes. I. Introduction: The analysis of bird songs and their expression in graphic notation." Ibis 103a: 231-245.
- Thorpe, W. H. and Pilcher, P. M. 1958. "The nature and characteristics of sub-song." British Birds 51(12): 509-14.
- Thorpe, W.H. 1964. “Singing.” In A New Dictionary of Birds, ed A. Landsborough-Thomson, 739-750. London: Nelson.
- Vella, Richard. 2000. Musical Environments: A Manual for Listening, Improvising and Composing. Strawberry Hills, Australia: Currency Press.
- Weisman, Ron; Ratcliffe, Laurene; Johnsrude, Ingrid; and Hurly, T. Andrew. 1990. "Absolute and relative pitch production in the song of the black-capped chickadee." The Condor 92: 118-124.
- Wilbrecht, Linda and Nottebohm, Fernando. 2003. "Vocal learning in birds and humans." Mental Retardation and Developmental Disabilities Research Reviews 9: 135-148.
- Williams, George C. 1966. Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought. Princeton: Princeton University Press.
- Williams, Heather; Cynx, Jeffrey; and Nottebohm, Fernando. 1989. "Timbre control in zebra finch (Taeniopygia guttata) song syllables." Journal of Comparative Psychology 103(4): 366-380.
Top >
